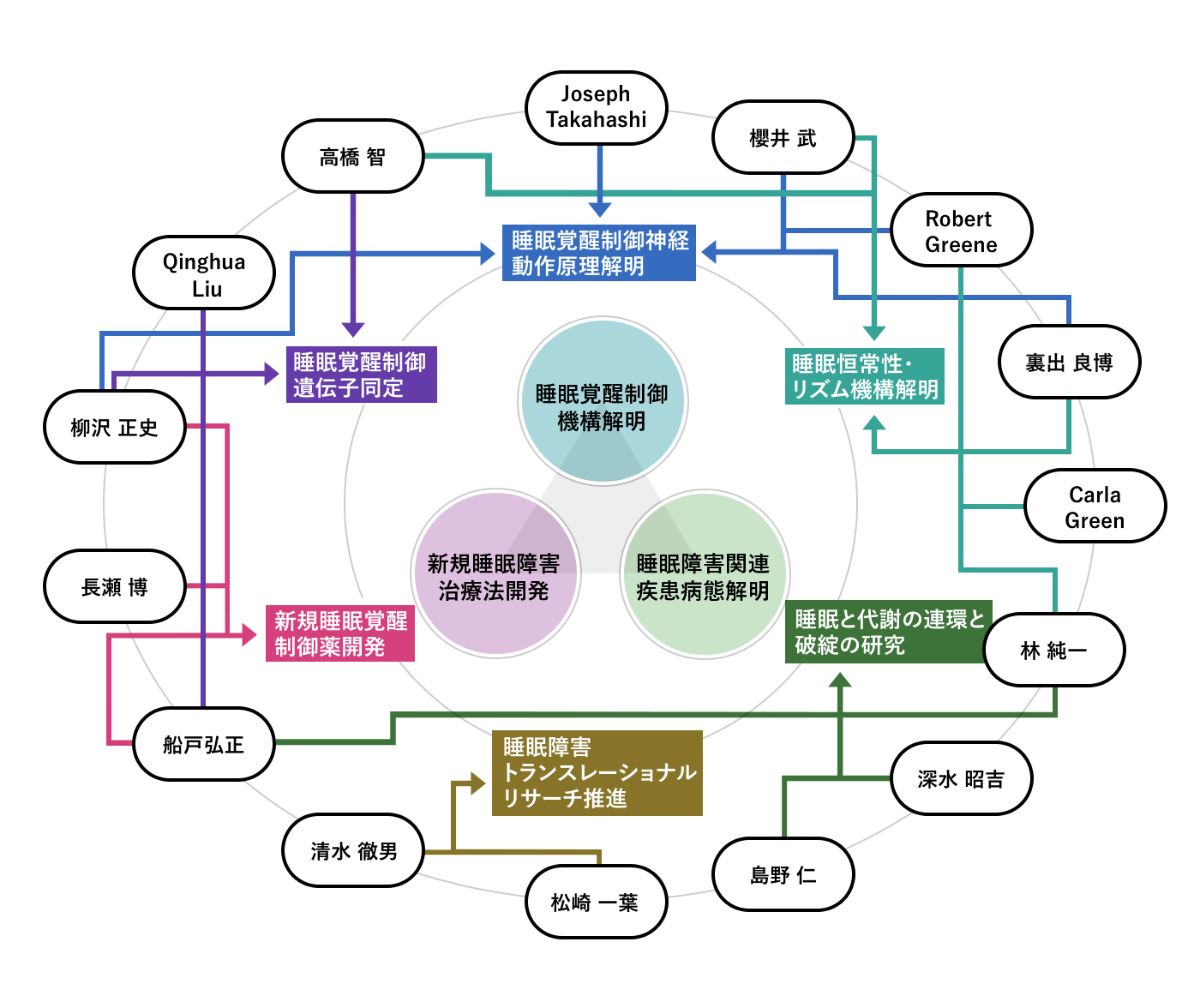
研究室について
IIISには現在20人の主任研究者からなる14のコア研究室と8つの連携・サテライト研究室があります。分子遺伝学・神経科学・生理学などの基礎生物学をはじめ、創薬科学、ヒト睡眠生理学など、多岐にわたる研究分野および国内外の研究者と有機的に連携することで革新的な睡眠研究を行っています。


睡眠覚醒の謎を解き明かし、人々が健やかに眠れる社会を作るための
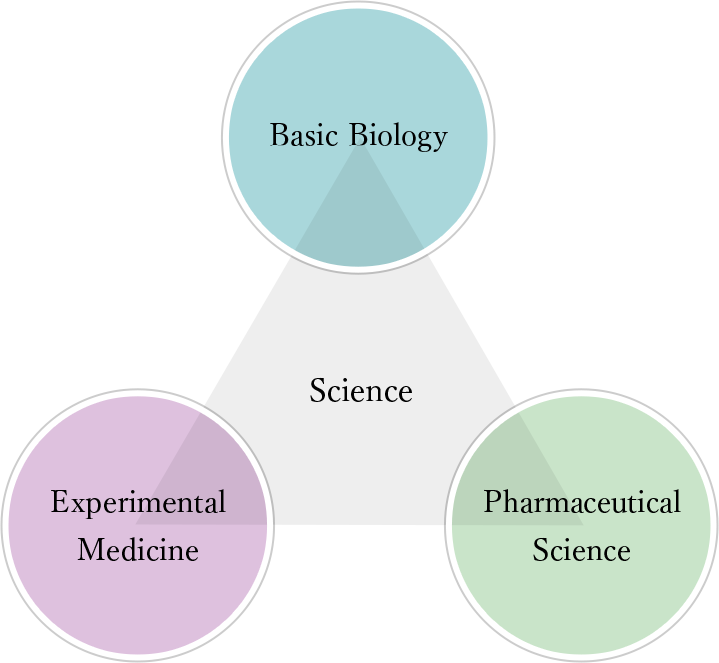
IIISは、「睡眠の機能と睡眠覚醒制御機構の解明」「睡眠障害と、それらに関連する疾患の病態の解明」および 「睡眠障害の予防法・診断法・治療法の開発」を3つのミッションとして掲げ、研究活動を行なっています。これらのミッションを達成するため、「神経科学」、「実験医学」、「創薬科学」という既存の研究領域を融合し、いわば睡眠の総合的なライフサイエンスである「睡眠医科学」という新領域を確立しました。
IIISには現在20人の主任研究者からなる14のコア研究室と8つの連携・サテライト研究室があります。分子遺伝学・神経科学・生理学などの基礎生物学をはじめ、創薬科学、ヒト睡眠生理学など、多岐にわたる研究分野および国内外の研究者と有機的に連携することで革新的な睡眠研究を行っています。

